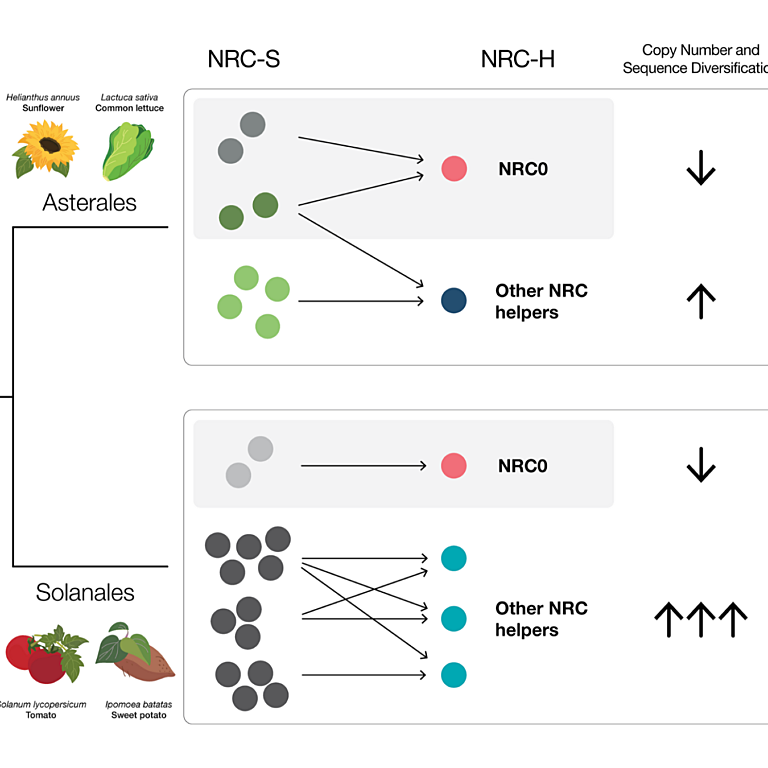
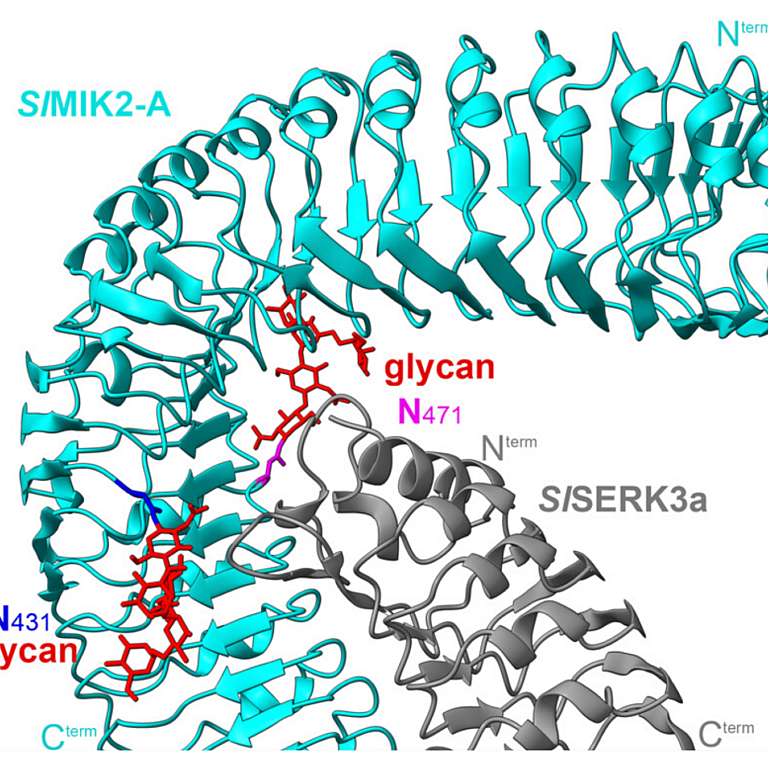
Oligomerization of a plant helper NLR requires cell-surface and intracellular immune receptor activation
Plant disease resistance involves both detection of microbial molecular patterns by cell-surface pattern recognition receptors and detection of pathogen effectors by intracellular NLR immune receptors. NLRs are classified as sensor NLRs, involved in effector detection, or helper NLRs required for sensor NLR signaling. TIR-domain-containing sensor NLRs (TNLs) require helper NLRs NRG1 and ADR1 for resistance, and helper NLR activation of defense requires the lipase-domain proteins EDS1, SAG101, and PAD4. Previously, we found that NRG1 associates with EDS1 and SAG101 in a TNL activation-dependent manner [X. Sun et al., Nat. Commun. 12, 3335 (2021)]. We report here how the helper NLR NRG1 associates with itself and with EDS1 and SAG101 during TNL-initiated immunity. Full immunity requires coactivation and mutual potentiation of cell-surface and intracellular immune receptor-initiated signaling [B. P. M. Ngou, H.-K. Ahn, P. Ding, J. D. G. Jones, Nature 592, 110-115 (2021), M. Yuan et al., Nature 592, 105-109 (2021)]. We find that while activation of TNLs is sufficient to promote NRG1-EDS1-SAG101 interaction, the formation of an oligomeric NRG1-EDS1-SAG101 resistosome requires the additional coactivation of cell-surface receptor-initiated defense. These data suggest that NRG1-EDS1-SAG101 resistosome formation in vivo is part of the mechanism that links intracellular and cell-surface receptor signaling pathways.